|
演化祕密的鑰匙?──淺談箕型門齒
文‧圖提供/邊鈺皓
在本館電子報《牙齒知多少》這篇文章,我們看到了烏山頭遺址的先民有著高比例的「箕形門齒」特徵。除此之外,這種舌面內凹的門齒特徵還廣泛出現在亞洲地區。對於這樣一個與特定群體有著顯著關係的門齒特徵,早在一百多年以前就有學者把它當作是解開族群祕密的重要鑰匙而受到關注。
 |
| Hrdlička(1920)原文圖板,圖中呈現了不同程度的箕形門齒。(Hrdlička(1920),Pl. I) |
箕形門齒與族群研究
體質人類學研究的重要學者Hrdlička在1920年以一篇「鏟形門齒(Shovel-shaped incisor,亦翻作箕形門齒)」(圖一)首次完整地針對這個普遍出現在北美原住民門齒的舌面內凹特徵進行命名與研究。除此之外,Hrdlička認為箕形門齒這個特徵與力學結構有關,基於演化上的緣由在白人群體中較為罕見,而在較為「原始的族群」中這一個特徵則較為普遍(註1)。
隨著後續學界對於「族群」有了更深入的探討與研究,並且納入了基因的概念,人體各類「非測量性特徵」作為多基因性狀,開始用以理解族群間關係及族群結構、歷史以及個體族群辨識的方法。而牙齒作為人體中最堅硬的部分之一,經常有著較為完整的保存,故而受到相當大的研究關注。箕形門齒這個特徵也因為學界研究發展有了更深入的探討,而再度受到大量的關注。
除了更全面性的研究發現,箕形門齒多見於亞洲及北美印第安人相關群體,其中又以北亞、北美印地安人的箕形程度為尤(Scott and Irish 2017)以外,由於許多古人類化石中也可以找到箕形門齒這個特徵,因此在人類演化關係的早期研究上,箕形門齒也有著相當重要的研究意義(Weidenreich 1937, Wolpoff 1989, Hanihara and Ishida 2005)。
箕形門齒的來源
有關箕形門齒發生的原因目前尚屬未明,現階段基因學的研究發現外異蛋白A受體(Ectodysplasin A receptor, EDAR)基因的變異似乎和這個特徵有所相關。而其中一個廣泛發現於亞洲及北美原住民族群,在rs3827760位點的小變異(註2)引起了大家的關注(Fondevila et al.2013)。
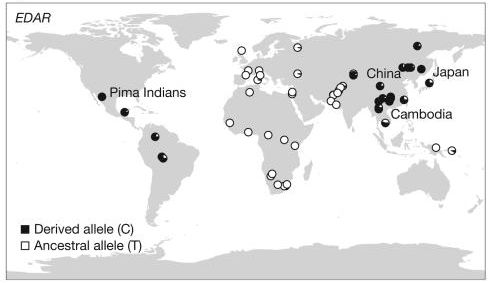 |
| EDAR V370A變異分布圖(引用自Sabeti et al.2007,Figure 2)。 |
從胚胎發育的角度來看,EDAR基因主要作用於胚胎的外胚層組織發育,所控制的部分除了牙齒以外,還包含皮膚、毛髮、指甲、汗腺等(NCBI, GeneID:10913)。當其中rs3827760位點的胸腺嘧啶(T)變異成胞嘧啶(C),其所對應的胺基酸也由由纈胺酸(valine,GTN)改變為丙胺酸(alanine,GCN)。根據研究(Fujimoto et al. 2008, Kimura et al. 2009, Park et al. 2012, Carayon et al. 2019),這樣的變異目前可以與箕形門齒特徵以及頭髮粗細及自然捲(註3)有所關連。除此之外其他特徵諸如下巴及耳垂形狀等性狀,似乎也與這個基因的變異有所關聯(Adhikari et al. 2016, Peng et al. 2016)。
綜上所述,箕形門齒在亞洲及北美原住民族群之間的普遍性,很可能並非是由於「箕形」這個特徵所帶來的好處。部分研究認為,這個在數萬年前開始興起於北東亞地區的特徵所產生的生存優勢源自於汗腺。進一步來說,在32000-15000年前,東亞地區因為季風的因素,相對其他地區而言較為潮濕,因此較為發達的汗腺可能具有較大的優勢(Kamberov et al.2013)。
另外有學者認為,這個基因的興起應該在更北邊一點的地方,由於在乾冷環境下,EDAR變異所產生的皮脂腺(特別是瞼板腺)增大可以延緩表皮水分的蒸散,起到潤滑的作用(Chang et al. 2009)。較為近期的研究則認為,EDAR的發展可能是因乳腺性狀的優勢,透過更為密集的乳腺,在兩萬多年以前白令海峽的環境下,寶寶得以獲得更為充足的維生素D,進而產生生存優勢(Hlusko et al.2018)。
以上研究雖然對於EDAR基因為何盛行於亞洲及北美印地安族群有不同的解釋,但都不約而同的同意EDAR基因是個複雜卻又隱含了人類演化、遷徙祕密的重要基因,其盛行背後所代表的很可能不是上述這樣單一性狀,而是由多性狀、多因素綜合起來的演化優勢。因此,現階段的研究僅能解釋部分的遺傳演化問題,後續尚待更進一步的研究。
人骨上的箕形門齒
如前述,有關箕形門齒背後的基因機制極為複雜,但如果我們從表現型的角度著手,箕形門齒實質上可能應屬於兩種特徵(遠側脊與近側脊突起)的結合,只是因這兩者的出現具有高度相關性而被當成是同一性狀(Scott and Turner 1997)。此外,除上顎前、側門齒外,下顎前、側門齒及上下顎犬齒亦可能具有「箕形」特徵,部分研究(e.g. Suzuki & Sakai 1966)顯示其各自的發生率略有不同。然而Turner II et al.(1991)則認為不同齒牙間箕形門齒的出現具有高度相關性,因此僅需考慮整體口腔中是否有「箕形」特徵出現即可。
針對上述觀點,筆者觀察記錄了東亞地區共248個個體齒牙的「箕形」特徵的出現與程度,單就上顎正中、側門齒來看,其箕形特徵的確在統計上具有相當程度的共現性,也就是說當任一顆齒牙具有「箕形」特徵,則另外一顆往往也具有「箕形」特徵。這個結果大致與目前基因研究成果之推論一致。然而,在觀察之中尚有少量(<10%)樣本具有左右不對稱且程度等級差超過2級的現象,其原因尚待解釋。
考量箕形門齒本身也是一個多基因性狀,雖然就目前為止我們發現了EDAR與他的關聯,但其背後尚有其他基因以及複雜的調控機制促使這個特徵的出現以及表現狀態。自百餘年前Hrdlička開啟了「箕形門齒」研究的鑰匙,目前的遺傳學及族群特徵分析研究已提供了我們一些初步線索來探究人類演化以及族群遷徙。隨著更大量的樣本分析、比較,我們對於世界各地區人群的分布、遷徙等也會有更深入的了解。
 |
 |
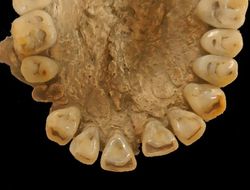
| 具「箕形」特徵之上顎正中門齒、側門齒。 |
不具「箕形」特徵齒牙。 |
 |
 |
| ASUDAS標準模板(左)箕形門齒-上顎正中門齒;(右)箕形門齒-上顎側門齒。 |
(註1)在Hrdlička的年代中,人們對於「族群」概念與現在有很大的不同。在當時多數人還具有「種族」以及「進化」和「原始」這樣的想法,因此本文特別加上引號表示為Hrdlička的想法。
(註2)在EDAR基因rs3827760點位所產生的SNP可被稱為EDAR T1540C(根據其核甘酸),V370A、370A、Val370Ala(根據其胺基酸)。
(註3)這裡所指的自然捲,是指東亞地區較為常見的那種頭髮捲曲,與另外一種歐美地區常見的頭髮捲曲方式有點不太一樣。有關歐美地區常見的頭髮捲曲,依照目前的研究,TCHH基因(Medland et al.2009)可能是其中一個重要的影響。
參考資料
Hrdlička, A. (1920). Shovel-shaped teeth. Am J Phys Anthropol 3(4), 429-465.
Scott, G. R., & Irish, J. D. (2017). Human Tooth Crown and Root Morphology. Cambridge, UK: Cambridge University Press.
Weidenreich, F. (1937). The dentition of Sinanthropus pekinensis: A comparative odontography of the hominids. Palaeontolagia Sinica, New Series D No. 101, No. 1.
Wolpoff, M.H. (1989). Multiregional evolution: The fossil alternative to Eden. In The Human Revolution, C.B. Stringer, ed. dinburgh, UK: Edinburgh University Press.
Hanihara, T., and Ishida, H. (2005). Metric dental variation of major human populations. Am J Phys Anthropol 128, 287– 298.
Fondevila, M., Phillips, C., Santos, C., Freire Aradas, A., Vallone, P. M., Butler, J. M., Lareu, M. V., Carracedo, A. (2013). Revision of the SNP for ID 34-plex forensic ancestry test: Assay enhancements, standard reference sample genotypes and extended population studies. Forensic Sci Int Genet 7(1), 63-74.
Sabeti, P. C. , Varilly, P., Fry, B.,(…). (2007) Genome-wide detection and characterization of positive selection in human populations. Nature 449(7164), 913-918.
National Center for Biotechnology Information (NCIB). (2004). GeneID 10913. Bethesda,MD: National Library of Medicine (US). [cited 2023 09 12]
Fujimoto, A., Kimura, R., Ohashi, J.,(…). (2008). A scan for genetic determinants of human hair morphology: EDAR is associated with Asian hair thickness. Hum Mol Genet. 17(6), 835-843.
Kimura, R., Yamaguchi, T., Takeda, M., (…). (2009). A common variation in EDAR is a genetic determinant of shovel-shaped incisors. Am J Hum Genet 85(4), 528-535.
Park, J. H., Yamaguchi, T., Watanabe, C., (…). (2012). Effects of an Asian-specific nonsynonymous EDAR variant on multiple dental traits. J Hum Genet. 57(8), 508-514.
Carayon, D., Adhikari, K., Monsarrat, P., (…). (2019). A geometric morphometric approach to the study of variation of shovel-shaped incisors. Am J Phys Anthropol 168(1), 229-241.
Adhikari, K., Fuentes-Guajardo, M., Quinto-Sánchez, M.,(…). (2016). A genome-wide association scan implicates DCHS2, RUNX2, GLI3, PAX1 and EDAR in human facial variation. Nature Communications 7, 11616.
Peng, Q., Li, J., Tan, J.,(…).(2016). EDARV370A associated facial characteristics in Uyghur population revealing further pleiotropic effects. Hum Genet. 135(1), 99-108.
Kamberov, Y. G., Wang, S., Tan, J., (…). (2013). Modeling recent human evolution in mice by expression of a selected EDAR variant. Cell 152(4), 691-702.
Chang, S. H., Jobling, S., Brennan, K., and Headon, D. J. (2009). Enhanced Edar signalling has pleiotropic effects on craniofacial and cutaneous glands. PLoS One 26;4(10), e7591.
Hlusko, L. J., Carlson, J.P., Chaplin, G.,(…). (2018).Environmental selection during the last ice age on the mother-to-infant transmission of vitamin D and fatty acids through breast milk. Proc Natl Acad Sci USA 115(19), E4426-E4432.
Scott, G. R., & Turner, C. G. (1997). The anthropology of modern human teeth: Dental morphology and its variation in recent human populations (2nd ed.). Cambridge, UK: Cambridge University Press.
Suzuki, M., & Sakai, T. (1966). Morphological Analysis of the Shovel-shaped Teeth. J. Anthropol. Soc. Jpn. 74(5), 202-218.
TurnerII, C. G., Nichol, C. R., and Scott, G. R. (1991). Scoring Procedures for Key Morphological Traits of the Permenent Dentition: The Arizona State University Dental Anthropology System. In Advances in Dental Anthropology, M. A. Kelley and C. S. Larsen, eds, NY: Wiley-Liss, Inc.
Medland, S. E., Nyholt, D. R., Painter, J. N., (…). (2009). Common variants in the trichohyalin gene are associated with straight hair in Europeans. Am J Hum Genet. 85(5), 750-755.
(本文作者為史前館南科館管理中心研究助理)
▲TOP |
|